Cấu trúc
Hêlicaza có nhiều loại, cấu trúc rất đa dạng và phức tạp. Thêm vào đó, mỗi loại hêlicaza khi chưa được kích hoạt và khi đang hoạt động luôn có cấu trúc khác nhau. Nhờ nỗ lực của nhiều nhà nghiên cứu, người ta thu được những hiểu biết ngày càng nhiều, mà trong mục này chỉ giới thiệu ở mức đơn giản.
- Mỗi hêlicaza khi hoạt động, thường ở trạng thái đa hợp phân tử (oligomeric), có thể ở dạng đồng hợp phân tử (homomeric) hoặc dị hợp phân tử (heteromeric), luôn liên kết với NTP (thường là ATP) và còn có thể liên kết với các chất khác.
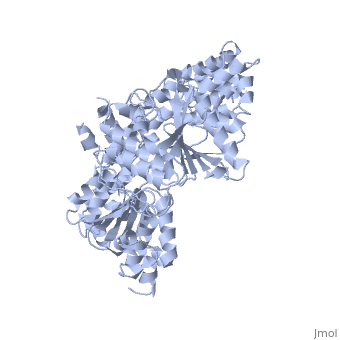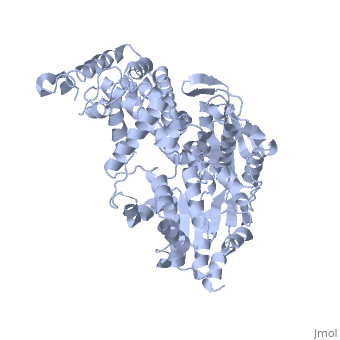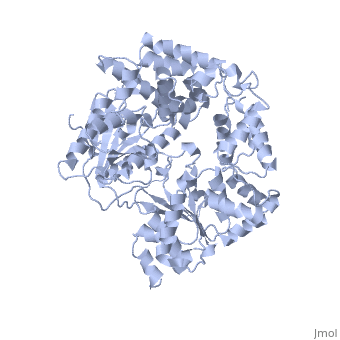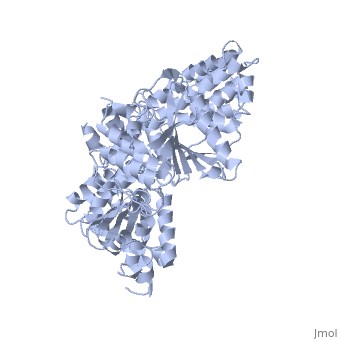
- Chẳng hạn, hêlicaza ADN B (dnaB helicase) ở vi khuẩn E.coli thuộc nhóm mã 1JWE và nhiều loại tương tự là tập hợp 6 phân tử (hexameric tức lục hợp phân tử) như biểu diễn ở hình 2, trong đó, mỗi phân tử là một tiểu đơn vị. Sáu tiểu đơn vị này tạo thành một cấu trúc hình vòng và được ổn định nhờ ràng buộc với NTP (GTP, CTP, TTP và thường là ATP) và có thể với ion kim loại, kết nối với cơ chất của nó (là axit nuclêic). Hầu hết các dữ liệu nghiên cứu thu được đều phù hợp với "mô hình vòng" này và cho rằng tập hợp lục hợp phân tử này tạo thành một cái "nhẫn" tí hon (helicase ring), bao quanh axit nuclêic mà nó xúc tác, vừa làm làm giảm khả năng nó bị rơi khỏi cơ chất, lại vừa giúp nó chuyển vị đúng hướng là "chạy" dọc chuỗi pôlynuclêôtit mà nó "ôm". Cái "nhẫn" này vừa ôm, vừa trượt dọc trên sợi ADN mục tiêu của nó (hình 3).[7], [8] Hầu hết hêlicaza có trạng thái đa hợp phân tử (oligomeric) như vậy, qua quá trình gọi là đa hợp phân tử hoá (oligomerization).
- Ưu điểm khác của kiểu cấu trúc hình nhẫn này là chu kỳ NTPase (như ATP-aza) có thể làm tăng hiệu quả và thúc đẩy nó "chạy".Tuy nhiên không phải hêlicaza nào cũng phải là đa hợp phân tử (oligomers) hình nhẫn. Nhiều enzym khác đã được chứng minh là hoạt động như là đơn phân tử (monomer) hoặc nhị hợp phân tử (dimers), chẳng hạn như RecBCD. Ngoài ra, nhiều nghiên cứu đã chỉ ra rằng hêlicaza có thể hoạt động thụ động hoặc chủ động tuỳ thuộc cấu hình của nó, cơ chất (phân tử đích) và môi trường.
Hình 2: Đồ hoạ giới thiệu cấu trúc một loại lục hợp phân tử (hexamers 1g8y).
Hình 3: Hêlicaza hình nhẫn đang hoạt động trên một mạch ADN.
Hình 4: Cấu trúc (đồ hoạ) của hêlicaza RuvA ở E. coli.
Hình 5: Hộp DEAD ở hêlicaza ARN.
Hình 6: Mô hình một hộp DEAD ở hêlicaza của người.
Xem mô hình 3D của một loại hêlicaza ở đây:https://proteopedia.org/wiki/images/b/b6/1pjr.gif
Cơ chế
- Hầu hết các hêlicaza cần một vùng axit nuclêic đơn để liên kết và bắt đầu hành động tách sợi. Theo cách diễn đạt nôm na, thì đó là một "đầu mẩu" của mạch đơn ADN hoặc ARN thò ra ngoài phân tử. Sau khi đã "bám" vào "đầu mẩu" này, nó dịch chuyển dọc sợi theo chiều 5′ –3 ′ hoặc 3′ –5′ (tuỳ loại hêlicaza). Như RecBCD chứa 2 tiểu đơn vị hêlicaza gần đối diện nhau nên có thể bắt đầu cả hai chiều. Tuy nhiên, hêlicaza hình nhẫn lại yêu cầu cấu trúc ADN hình chữ Y (ở chạc nhân đôi) với một sợi cơ chất và một sợi không bổ sung có độ dài tối ưu để bắt đầu dãn xoắn và cắt liên kết hyđrô.
- Còn có loại hêlicaza tương tác cùng lúc với cả hai sợi và di chuyển theo một chiều, hoặc cả hai chiều.[9]
- Trong hoạt động, hêlicaza là một động cơ phân tử lấy năng lượng hóa học từ phân giải NTP để thủy phân tách các chuỗi bổ sung của các mạch kép, đồng thời cũng loại bỏ các prôtêin "bám" lên cơ chất, tạo điều kiên cho tự sao mã, phiên mã hoặc xúc tác tái tổ hợp tương đồng giữa các ADN.
- Hoạt động enzym của hêlicaza làm dãn xoắn axit nuclêic được thực hiện thông qua việc hạ thấp hàng rào kích hoạt B của mỗi hành động cụ thể. Hàng rào này là kết quả của nhiều yếu tố khác nhau và có thể được xác định bằng cách sử dụng phương trình sau, trong đó:
N {\displaystyle N} = số cặp bazơ cần dãn xoắn (bp),
Δ G b p {\displaystyle \Delta G_{bp}} = năng lượng cần để tạo cặp bazơ,
G i n t {\displaystyle G_{int}} = mức giảm năng lượng tự do nhờ hêlicaza,
G f {\displaystyle G_{f}} = mức giảm năng lượng tự do nhờ tháo xoắn.
B = N ( Δ G b p − G i n t − G f ) {\displaystyle B=N(\Delta G_{bp}-G_{int}-G_{f})}
Các yếu tố góp phần vào chiều cao của rào cản kích hoạt bao gồm: trình tự axit nuclêic cụ thể của phân tử liên quan, số lượng cặp base liên quan, sức căng hiện diện trên nhánh nhân bản và lực bất ổn.
- Kích thước của rào cản kích hoạt để khắc phục bởi hêlicaza góp phần vào việc phân loại nó là hêlicaza chủ động hoặc thụ động. Trong nhóm hêlicaza thụ động, một rào cản kích hoạt đáng kể tồn tại (được định nghĩa là B>kBT; trong đó kB là hằng số Boltzmann và T là nhiệt độ của hệ thống). Bởi vì rào cản kích hoạt này là đáng kể, nên tiến triển tác động của nó bị ảnh hưởng chủ yếu bởi chuỗi axit nuclêic trong phân tử đích cần giải phóng (cần cắt), và sự có mặt của các yếu tố khác lên chạc nhân đôi (chạc chữ Y[4]). Một số loại bazơ ở axit nuclêic có thể làm giảm tốc độ dãn xoắn và tách mạch, như G (guanin) và X (cytosine) do có số liên kết hyđrô nhiều hơn A với T, ngược lại một số yếu tố khác lại có thể tăng tốc độ này. Trong nhóm hêlicaza, thì tốc độ dãn xoắn và tách mạch Vun nhỏ hơn tỷ lệ chuyển dịch Vtrans trên sợi đơn.[10]
- Trong nhóm hêlicaza chủ động, thì B<kBT, do rào cản ít đáng kể, vì hêlicaza có thể làm mất ổn định axit nuclêic,tăng tốc độ dãn xoắn và tách mạch, bất kể chuỗi axit nuclêic nào. Ở nhóm này, Vun = Vtrans. Nhóm này còn có khả năng trực tiếp gây mất ổn định chạc nhân đôi để thúc đẩy dãn xoắn và tách mạch.[10]
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
